
I had a brief conversation on BioLogos with someone who’d been a little troubled by the ancient arguments about “vestigial organs”, found in Darwin and echoed in textbooks for kids since as proof of evolution. I ended up suggesting that in very concept the idea is worthless and ought to be dropped. The trouble is that, like much of the popular presentation of evolution, it is actually not a good argument for evolution, but a polemic “case for evolution against creation”, and so is ideologically driven, even though it’s of no value to science and less to theological discourse on creation.
Conor Cunningham shows how much more complicated the actual state of science and the state of theology is than that, and uses the term “Vulgar Darwinism” for the simplistic version of Darwinian evolution that gets to the public. Unfortunately this also dominates a significant subset of those in the scientific profession itself, and for their creed Cunningham uses “Ultra Darwinism” as a synonym. We all know who some of the professional perpetrators are, for they include some of those most involved in popularising evolution and fundamentalist materialism.
To cover “vestigial organs” briefly, then, they are a subset of homology, and homology is undoubtedly a good strand of evidence for common descent. However, even in that restricted use homology is inconclusive evidence for Darwinian natural selection. I’ve written before that to Linnaeus, the father of taxonomy before evolution, homology supported the “plenitude” principle that God creates every possible variation.
And if we ignore that, there is increasing evidence of some kind of formal constraints on “design” in nature (structuralism), one aspect being convergent evolution, and that is probably a better explanation for homology that any version of Darwinian theory.
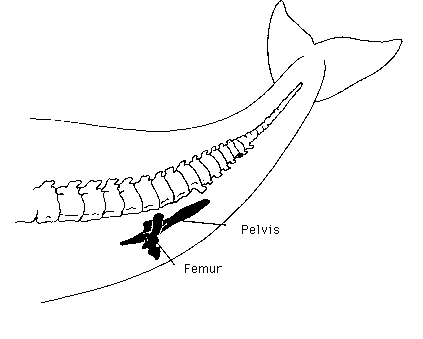
The real problem with “vestiges”, though, is that in evolutionary terms it’s a merely subjective judgement to say what is a “normal” function, and what a “vestigial” one. For it’s rare to find organs that can be shown to be truly functionless. The human appendix has important immune functions. It’s commonly said that it’s a vestigial version of the large caecum of, say, rabbits, that enables the digestion of cellulose.
The common argument that the appendix is unimportant because it can be surgically removed without harm is weakened considerably when one discovers that the same is true of the rabbit’s caecum. This was done as long ago as 1955, and the researchers were able to say:
Successful operations have been performed on 5 rabbits to
date, and these have lived for extended periods, up to 9 months after cecectomy, with no external evidence to indicate that growth and development had not been entirely normal.
Furthermore who is to say that the rabbit caecum itself is not an extreme specialisation of another role, even a primary immune function like that of the appendix? Many evolutionists believe that virtually every organ’s function is an exaptation of some other use in any case: dinosaur insulating feathers become flight surfaces, fins can become legs and so on.
 Within Darwinian science, vestigial organs really only provides a weak and subjective way of judging between the adaptationist and neutralist camps: adaptationists stress how natural selection ensures that everything is useful, whereas neutralists say that the limits of selection mean that most evolution happens by chance. Only the latter really ought to believe in vestigialism – the adaptationists ought to jump on any existing function as an adaptation, or claim we’re bound to discover it if we look hard enough (which we usually do).
Within Darwinian science, vestigial organs really only provides a weak and subjective way of judging between the adaptationist and neutralist camps: adaptationists stress how natural selection ensures that everything is useful, whereas neutralists say that the limits of selection mean that most evolution happens by chance. Only the latter really ought to believe in vestigialism – the adaptationists ought to jump on any existing function as an adaptation, or claim we’re bound to discover it if we look hard enough (which we usually do).
But in practice, both groups, and those in between, tend to use vestigial organs only as one of those ubiquitous theological arguments for evolution against creation. “Look, a sensible God wouldn’t make such useless wisdom teeth/coccyges/back limbs on whales and so on. Ergo evolution is true.” But of course, the judgements about what is or is not “function” remain every bit as subjective as before, only by mentioning God they have invoked all kinds of possible final causes beyond mere function, from aesthetics to deliberate curtailment of fitness for the general benefit of creation. Remember, “biological fitness” as a be-all-and-end-all is a strictly evolutionary concept in which theology has no great stake.
So I suggest the whole concept of vestigial organs achieves nothing, and doesn’t even deserve attention from the Christian either for or against evolution, since for them the reality, but inscrutibility, of God’s purposes ought to be axiomatic.
Not long after that BioLogos conversation, I remembered a comparable example of “vulgar evolution apologetics”: the so-called “evolutionary dead-end”. Paradoxically I came across it last in Conor Cunningham’s book Darwin’s Pious Idea, which is all about the deeper philosophical implications of Darwin, but seemed to have accepted the particular category of “evolutionary failures” unchallenged, as incontrovertible fact.
But once again it’s a mainly theological concept with no real scientific value – “Look at all the waste in evolution, with all those failed experiments littering the field. Would a designing God really have shown such incompetence, especially given the consequent suffering for that majority of badly-designed types?” Cunningham uses the illustration of the lottery winner who thinks he’s lucky, failing to appreciate that he’s just the only non-loser among millions.
So evolutionary “dead ends” and “failures”, it is held, demonstrate the non-directedness of evolution. But the question is, what demonstrates the existence of such dead ends and failures? In fact, they are circularly assumed to exist from the predictions of the Darwinian model of evolution. It’s the same argument that says the fossil record only contains 1%, or whatever, of all the types that have ever existed, as support for gradualism, when gradualism itself was used to claim the incompleteness of the fossil record – see here.
Extinctions are often cited as direct evidence of these “evolutionary dead ends” as if the connection were self-evident. But extinctions do not demonstrate failed directions of evolution, any more than the death of soldiers in battle demonstrates failed strategies. Rather they show changes of conditions that eventually exceed that type of organism’s potential for adaptation. Steam engines were not dead-end technologies because they were replaced with diesels. It only becomes a “failed experiment” if you make the continuance of a particular line the sole criterion of success, rather than (say) the health of the whole biosphere – which is more likely to be God’s concern.
It only becomes a tragic failure if you weave some fanciful image of the Last Fish dying slowly in a desert with gasping regrets she’ll never see her grandchildren – a scenario which in real life would be very much like the millions of other similar fish that died in deserts fortuitously during the millions of years it was a flourishing species. Why is extinction any more wasteful and tragic than death? Both bequeath the earth to other living individuals
So extinctions as such count for nothing as evolutionary mistakes. What we’re looking for are the abject failures: not the steam engines that gave way to diesels, but the steam toasters or steam battery packs that some moron patented before they sank into welcome oblivion. And those are harder to come by in the biological world. Conor Cunningham’s lottery analogy is only valid for lotteries because one meets many lottery losers in life – both losers and winners are available for interview.
But the fossil record actually shows an overwhelming predominance of apparent stasis and sudden changes of (successful) forms (hence punctuated equilibria). So gradual transitions, even in the more rapid punk eek sense, are in pretty well all cases postulates of the assumption that they “must” have existed, but not fossilised. The same is true of “evolutionary failures” – they may have occurred during punctuated equilibria, or even as isolated lineages during stasis – but they don’t actually manifest as fossils. They are a pure hypothesis.
 How would we even tell such a “failed experiment” if we did find it? It would represent some species that once lived, and a live species is, by definition, more or less successful. Stillborn fetuses seldom fossilize – and never take part in evolutionary experiments. Low or declining numbers might be because the species is a failure – but more likely are demonstrably because the species occupies a specialised niche that disappears. Or it may even just lose out in a common role due to exceptional circumstances (eg migrating grey squirrels ousting reds in the UK, global warming leaving nowhere for cold-loving species to go, etc).
How would we even tell such a “failed experiment” if we did find it? It would represent some species that once lived, and a live species is, by definition, more or less successful. Stillborn fetuses seldom fossilize – and never take part in evolutionary experiments. Low or declining numbers might be because the species is a failure – but more likely are demonstrably because the species occupies a specialised niche that disappears. Or it may even just lose out in a common role due to exceptional circumstances (eg migrating grey squirrels ousting reds in the UK, global warming leaving nowhere for cold-loving species to go, etc).
Some have said the giant panda ought to be quietly left to go extinct extinct because it is so poorly adapted. But it’s actually been successfully around since the Pleistocene, which is as long as we have. If its habitat or other environmental factors are no longer conducive to its survival, then that’s another matter. The dead end was not due to evolutionary incompetence, but to deforestation and so on.
 How would one tell any fossil is a “dead end”? Extreme bauplans are no guide. We can have little idea of the entire world a strange creature lived in, so it’s impossible to say how well suited it was. It’s even impossible to be sure how rare it was, because by all accounts the fossil record is patchy. After all plenty of today’s common plants and animals are weird.
How would one tell any fossil is a “dead end”? Extreme bauplans are no guide. We can have little idea of the entire world a strange creature lived in, so it’s impossible to say how well suited it was. It’s even impossible to be sure how rare it was, because by all accounts the fossil record is patchy. After all plenty of today’s common plants and animals are weird.
For example, commonly it was said (and still is, in popular science texts) that the Irish elk became extinct because its increasingly disproportionate antlers, assumed to be sexually selected, eventually made the whole creature maladaptive. But all one really knows is that they had big antlers and went extinct, and the cause is an hypothesis – and one that doesn’t bear much close  examination.
examination.
The redoubtable iconoclast Stephen J Gould drew attention to findings that as deer increase in size, their antlers customarily enlarge disproportionately. The giant elk, being the largest deer found, had antlers that followed this rule. This suggests that some kind of developmental constraint, or “law”, was probably at work in the elk, rather than any evolutionary “experiment”, whether that experiment were entirely directionless, an attempt to adapt to some supposed need that failed, or even the inexplicable love of the elk ladies for grotesque headgear.
But we don’t really need Gould’s excellent analysis to show the vacuity of the “dead end” hypothesis. For in fact Megaloceros giganteus (the giant elk’s posh name) shows in the fossil record from 400,000BC to 8000BC. That longevity in itself undermines any idea of a failed experiment. More significantly still, there seems to be no evidence that their antlers actually increased in size over that time – otherwise they’d be the missing example of gradualism that paleontology needed so badly to keep its place at the classical Darwinian table last century. The real argument, it seems, was simply, “Those antlers look too big to work. And the elk’s extinct. They must have evolved gradually because everything does. Therefore they just got too large and the experiment failed.”
It’s just another “Just So” story (that astute analogy was from Gould, too). In fact, it looks like giant elks died out because of the usual issues of lack of the right food when the ice-age ended, perhaps aided by human spears and arrows.
So, maybe evolution makes experiments that go nowhere, and maybe it doesn’t, for all that the evidence shows. And maybe God makes such failures for his own reasons, and maybe he doesn’t. But the arguments for and against either explanation would be stronger if the whole menagerie of such failures weren’t entirely subjective, or perhaps even entirely imaginary.
Great piece, Jon, and I totally agree with your points about “dead ends” and extinction as some of sign of moral ineptitude on God’s part. In fact, I am in quite the minority of environmentalists, since I do not necessarily view ALL extinctions as a bad thing by definition. For those who do, I tend to ask, “bad for whom?”
I also think that convergence is going to become a major focus for future research in evolution. It is no longer an interesting anomaly, but the possible basis for how biology works. Of course we have never seen the real failures in evolution, Conway Morris mentions wheels on animals, and I came up with jet flight as two things that never made it off the ground (sorry). There are no intrinsic biological rules against such things, (wheels, jets and wings all exist) but the law of convergence states first and most clearly “Everything does NOT go”. What could be exciting would be find out if we can find out exactly what kinds of constraints operate (on a molecular level) and what that means for evolutionary direction, and perhaps, teleology.
Good morning Sy (at least, it’s morning here!)
Do you think one day a flying octopus may evolve to converge with flying fish, and then your jet flight example would have to be crossed off?
Conor Cunningham cites Conway Morris quite a lot (but sadly I missed hearing him when I got held up in traffic on the way to Cambridge a few years ago). I suppose my main quibble with him is that the emergent properties he seems to invoke for convergence remain entirely mysterious, yet he is rather confident in his assertions about their naturalness and so on, in disdainful opposition to what he deems foolish ID explanations.
Yet convergent evolution remains, it seems to me, at the purely descriptive stage – explanatory mechanisms are well down the line, and “emergence” seems to be a kind of magic wand to cover ignorance (rather like the famous “dormitative power” of opium example).
I therefore find his global “predictions” – eg that life is bound to produce something like man – rather gung-ho. We still only have one example of such a being – us – and we don’t know nearly enough to say how precise a “tool” convergence is. Perhaps it’s not just that streamlined forms like porpoises and ichthysaurs are inevitable, but that the actual reptile and mammal species with those forms are.
The exciting thing though, as you say, is that if one is forced to admit constraints on the open-endedness of evolution, and if one can elucidate the secondary powers that make it so, it would make the richness of the biosphere far more explicable than randomness-based theories, and would fit the fossil record better. Biology would become law-based.
Needless to say though, it would also have profound theological implications, because the very business of downgrading randomness upgrades teleology. If something hasn’t happened by chance, it’s happened by design, even if that design goes back to the basic structure of the universe.
That’s why I’m bemused that Conway Morris is so vehemently against ID on this: he’s uncovering a form of fine-tuning, which is in the IDists’ terms “design by frontloading”, and distinguishable from chance (or if not, why not?)
One more thing on this: I can appreciate that to do science, one is necessarily looking at the regularities that underlie reality. But theologically, if that leads one to insist on the universe as a closed causal system (essentially embracing Deistic explanations), one is forced to a theology that makes a virtue of God’s non-involvement in nature, and that is the opposite of the biblical picture (as my last few previous posts have tried to look at).
In practice wedding theology to science too much works out like this to TEs: if you’re Neodarwinian, evolution is open-ended, so God can’t have particularly willed mankind or individual species. It must be because of the Cross – he took the costly risk of giving the universe freedom and not exercising his power.
If you’re into emergence, though, God set up the universe so an intelligent species like us was inevitable, but didn’t have any opinion on how many fingers and such details – nor did he plan more than the general outcomes of species. Maybe we have to emphasise more of the wisdom of the Logos, and go a bit easier on the kenosis.
But in my view, a true Christian theistic science will study whatever secondary causes can do, however vague or precise, and yet allow the remainder to be outside science, but within providence. Whatever is not determined by secondary causes, in other words, is still under God’s governance in some other way that science cannot describe.
That way, God created not streamlined marine vertebrates, but Caypullisaurus, and not a generic intelligent being, but that wonderful girl with the quirky humour and beautiful eyes. He set up the ecological systems, but he actually feeds the young lions and ravens.
Does that seem anti-science to you? 🙂
I have served on an advisory committee with Simon, and you will not be surprised to learn that he has a mind that I can only hope to catch up to with a great deal of effort, time and thought. He also speaks exactly how he writes, so that can be a challenge as well for the more slow witted (like myself). I am in the process of reading his book Inevitable Humans in a Lonely Universe, which tends to make one breathless. A remarkable man. And a Christian, as Im sure you know. There are hints of this throughout the book, but he tends to be careful (as are we all) in a strictly scientific setting.
I fully agree with your comment about the need for more knowledge about how the whole convergence thing works, and by good fortune, I am actually working on that (related to gene regulatory networks). Andreas Wagner has some very interesting conclusions there, but I am interested in going a bit deeper and hopefully further.
As for whether convergence explains humans are just smart bipeds or Jon Garvey himself, I have no idea. I again completely agree that there is a great STOP sign along every scientific road, which in order to pass, we must dismount from our rational naturalism vehicles, and find a different kind of conveyance to take us further.
We have found those STOP signs in physics and math (thanks to Heisenberg and Godel) but most biologists doubt they exist for them. I dont, and I am eager to come across one. I think that such a finding would in itself be a pretty good indication (and an irrefutable one, like the uncertainty principle and the incompleteness theorum) that God must play a pretty active role in biology as well as in the rest of creation. Something we know by faith and Scripture already, but wouldnt it be nice to see it in our science.
Sy
“A man sees what he wants to see
And disregards the rest.” (Paul Simon)
I don’t doubt the stop signs are even more numerous in biology than in physics, as there are vastly more levels of improbability. But both here and at BioLogos one has heard atheists saying that the evidence that quantum events have no cause within nature disproves God as a cause.
Whether it is possible to show an active role for God in biology is a moot point – though it depends what one means by demonstration. If you read my post on Jesus’s prophecy over Jerusalem, I suggested that the events of 69-70AD, being very well documented historically, show no obvious explanatory gaps requiring God.
The evidence for God’s involvement in them comes simply from the spec ificity of Jesus’s prophecy, which can, of course, always be either denied or ignored as luck.
Likewise I think it will be the inevitability of teleology in biology, and the lack of decent materialist accounts of teleology (Thomas Nagel’s being a prime example) that may be all we can *see* of God in science.
I posed a thought experiment on BioLogos a long while ago in which a scientist (based on Lenski!) discovers his culture of E.coli has become an entirely different species overnight – in other words, a frank miracle. Even in that hallowed setting, there was at least on biologist saying that the true scientist would continue to seek for an explanation – chance, if necessary. Chance is, of course in the end formally indistinguishable from miracle.
I certainly wish you well on the gene network project, whilst considering ruefully that if you and your team even begin to get your heads round it, it will be quite incomprehensible to me. It’s the apex of non-reductionism, as far as I can see!
Are you implying I’m that wonderful girl with the quirky humour and the beautiful eyes?? It’s all lies, I tell you.
Thanks for all your interesting and thoughtful remarks here, Sy. I look forward to reading more of your comments on evolutionary mechanisms, convergence, the characteristic blind spots of biological science, etc. It’s good that you now feel freer to speak out here and on BioLogos.
Jon, do you see pseudogenes as real examples of vestigial structures? Especially in the cases where there are species with the functional form, and multiple other species with non-functional forms containing the same inactivating mutations, it seems like pretty plain evidence for common descent, ignoring any theological implications that anyone might assert.
Hi Preston.
I hadn’t been considering vestiges at the molecular scale (I’m still a frustrated palaeontologist at heart), but seeing as it’s the theological assertions I was arguing against, I’d agree that once again pseudogenes are evidence in favour of common descent.
Even so some of the same caveats applicable to “vestigial organs” might apply; for example, if there are law-like constraints on the kinds of molecular variations that occur, then identical sequences in similar species could have a non-Darwinian alternative explanation. I suppose the homologous proteins given as evidence for convergent evolution are part of that.
Introduce “any sensible God wouldn’t let genes break”, though, and one’s into dubious theology again: that applies whether one is considering God’s final causes or any teleological secondary causes (which, being imperfect, are bound to go wrong sometimes).
But overall, like homologous organs big and small, pseudogenes most plausibly imply evolutionary relationship. But that’s only worth thinking about if you’re trying to convert a Creationist, and one would think biologists had better things to do.
A PS to this – I’ve become aware of research that shows unique functions for some pseudogenes in that complicated genetic and epigenetic soup we call life.
If that’s true, it seems to put pseudogenes even more in the same category as vestigial organs – evidence for homology, or alternatively for convergence, because they’re similar across species and function differently (which means they’re not really similar at all).
Another PS – an interesting line on the appendix as a vestigial organ here. It turns out the appendix is apparently always an immune organ, and was never for digestion, even in the rabbit.