
That title is, of course, a gross exaggeration or indeed a calumny (but hey, it sounds good!): cladistics is a tool that is useful according to how it is used. But a major 2009 paper on the evolution of birds addresses some pitfalls in its common use, and points out that:
Cladistics should be treated not as a way to test phylogenetic hypotheses but as an exploratory method, useful, if handled sensitively, for comparing and evaluating hypotheses.
The paper is eight years old now, and from outside the field it is hard to evaluate whether the doubts this paper throws on the common story that the birds evolved from the maniraptorian theropod dinosaurs (which the authors term the “BMT hypothesis”), are still valid today. There are certainly new complexities to deal with since then.
My interest here is the authors’ examination of the methodology of cladistics, which is still often held up as slam-dunk evidence of nested hierarchies, and therefore of Darwinian evolution. I’ve shown before that nested hierarchies were taken as evidence of special creation long before they became evidence of evolution, and so they prove nothing about those as alternative theories of life. But the question here is how representative of reality those hierarchies are in the first place. The authors say that cladograms are often wrongly treated as evidence for particular phylogenies:
Even though a cladistic analysis does not actually test a hypothesis — it simply produces the most parsimonious phylogeny, given the assumptions incorporated into the analysis a priori – many have claimed that the BMT hypothesis has been tested with cladistics (e.g., Padian and Chiappe 1998; Padian 2001b; Prum 2002, 2003).
This is an example of how, indeed, theory governs observation, as we shall see – though not in a Postmodern Deconstructional way, but at an everyday level of assuming more than evidence warrants. In more (technical) detail they say:
The BMT hypothesis has been claimed to be overwhelmingly supported by the many synapomorphies shared by birds and theropods (e.g., Padian and Chiappe 1998; Chiappe and Dyke 2002; Paul 2002; Prum 2002, 2003; Xu et al. 2003; Xu 2006). The hypothesis is inferred to be correct because of the vast quantity of data held to support it. This argument is logically problematic. The number of confirmatory observations that a hypothesis can marshal is not necessarily relevant to whether the hypothesis is correct, and the history of science offers numerous examples of “overwhelmingly supported” theories that were later refuted (Hempel 1966; Popper 1998, 2002). The risk is that only supporting evidence will be recognized, while contradictory evidence is ignored or explained away. Kluge (1997, 1999, 2001) has repeatedly urged the adoption of a falsificationist, rather than a verificationist, research program for cladistics.
This is the old falsification issue long ago raised by Popper: piling on cladistic correlations to a single hypothesis, in this case the evolution of birds from theropods, actually proves nothing at all unless one can also deal (a) with alternative possible phylogenies and (b) with the evidence that contradicts the proposed clade, of which some always, in practice, exists.
This second point is easily illustrated by a classic nested hierarchy from our grand-parental toybox:
Get a series like this, and the family relationship looks intuitively very convincing. The problem is that in biology the series are never either that neat or complete. And there are always anomalies, which in the case of avian origins are quite serious.
Consider this second, smaller, nested hierarchy:
In terms of some features, such as general morphology and overall size gradation, we have a clade – which tends to break down as we add up the differences in detail. In this case, given our general knowledge, we know that this hierarchy exists only at the conceptual design level of “Russian Dolls”. In fact the middle doll is ontologically unrelated to the others – or involves convergence or some other auxiliary hypothesis to account for the anomalies. Our authors show that such “excuses” are common in the scientific literature:
Introduction of adhoc auxiliary hypotheses.—An ad-hoc auxiliary hypothesis is one that has been formulated for the specific purpose of restoring agreement between a hypothesis and falsifying observations; it serves no independent explanatory function and does not entail any significant, independently testable implications (e.g., Hempel 1966, Popper 2002). Although ad-hoc auxiliary hypotheses are often used to protect favored hypotheses (Kuhn 1970), and although they may be empirically valid, they actually interfere with testability (Hempel 1966, Popper 2002) by increasing the range of observations with which a hypothesis is compatible. If the introduction of ad-hoc auxiliary hypotheses were considered legitimate, they could be used to explain away all falsifying observations, rendering a favored hypothesis immune to any criticism. Repeatedly obtained observations that contradict a hypothesis should be accepted as falsifying observations rather than explained away.
The authors give an example which actually ought to be quite problematic for the theropod to bird hypothesis. The three-toed foot of theropods is embryologically derived from digits 1-3 of the primitive pentadactyl limb. Birds, however, derive theirs from digits 2-4:
This repeatedly obtained observation contradicts a key prediction of the BMT hypothesis and is logically sufficient to refute it. To explain away this falsifying observation, Wagner and Gauthier (1999) proposed that a homeotic frameshift affecting digital identity must have occurred during the evolution of neotetanurine theropods.
Now of course, this presto-changeo solution to explain the non-homology of supposedly homologous features may be true. But it is still explaining away inconvenient evidence to save a theory, and one can easily find other examples of this throughout the biological literature. For example, theropods also have a forward-pointing pubic bone. This feature formed the 130 year old fundamental division between them (the Saurischians) and the other main branch of dinosaurs, the Ornithischians, in which the pubis is tipped backwards – like birds, supposedly descended from saurischian theropods. But a paper published this year proposed a completely new dinosaur phylogeny based on the assumption that this major anatomical change “must have” evolved at least twice by convergent evolution (the modern Ornithoscelida hypothesis).
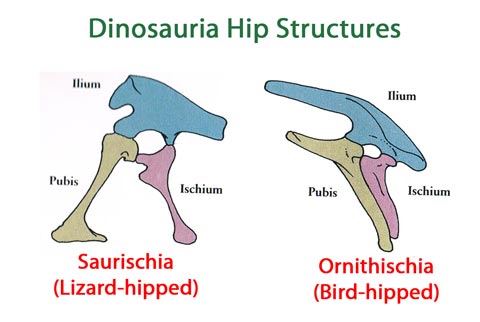
The two classic types of dinosaur hip – the problem is that the birds are believed to derive from saurischians, not ornithischians
This hypothesis is adopted only to bridge the anomalies in avian evolution, and taking it as an assumption will inevitably generate new cladograms, even if the assumption, or indeed the whole evolutionary construct, were mistaken. Recurrent evolution of the same feature “saves the appearances” for conventional evolutionary processes, just as gaps in the fossil record are routinely filled with hypothetical transitional forms, or the fossil record’s low “resolving power” is exploited to explain how punctuated equilibria can produce speciation by conventional mechanisms and yet be always invisible, because they are stated to be of too short a duration to be shown in the fossils.
To me, the most interesting part of this paper is its examination of the cladistic “case” for the BMT hypothesis. First they repeat the work by which the “verificationist” approach endorses the birds-from-theropods hypothesis, and agree with the closeness of the match:
We reanalyzed a standard matrix of 46 taxa and 208 characters from a recent paper by Clark, Norell, and Makovicky, and we found statistical support for the clades Coelurosauria and Maniraptoriformes and for a clade of birds and maniraptorans. Note, however, that because the matrix contains only birds and theropods, it assumes that the origin of birds lies within the Theropoda.
You see the argument here; since the theory is that birds evolved from theropods, only birds and theropods are examined. And lo and behold, cladistics confirms the theory, because methodologically it was bound to: the only point in question was the best fit within that assumption. But:
In addition to this problem, Clark et al.’s (2002) matrix contains problematic assumptions of homology, especially in the palate, basipterygoid, manus, carpus, and tarsus. In an attempt to avoid these two major problems and to evaluate the BMT hypothesis and four alternative hypotheses in a comparative phylogenetic framework, we followed the recommendations of Jenner, Kearney, and Rieppel by constructing and analyzing a larger but more conservative matrix. Our matrix includes taxa from throughout the Archosauria.
They add some technical procedural detail, of course, but the bottom line is that they have broadened their dataset to include the possibility that birds are descended from some other taxon, rather than maniraptors, including the hypothesis that the maniraptors, rather than being the ancestors of birds, are in fact flightless examples of birds. And the result was that either cladogram (and a couple of other unrelated ones too) was equally possible:
Kishino Hasegawa tests revealed no statistical difference between the hypothesis that birds were a clade nested within the Maniraptora and the hypothesis that core clades of Maniraptora were actually flying and flightless radiations within the clade bracketed by Archaeopteryx and modern birds (Aves). Additional statistical tests showed that both the “earlyarchosaur” and “crocodylomorph” hypotheses are at least as well supported as the BMT hypothesis. These results show that Theropoda as presently constituted may not be monophyletic and that the verificationist approach of the BMT literature may be producing misleading studies on the origin of birds. Further research should focus on whether some maniraptorans belong within Aves, and whether Aves belongs within Theropoda or is more closely related to another archosaurian taxon. At present, uncertainties about the hypothesis that birds are maniraptoran theropods are not receiving enough attention.
Now the point is that, whether or not the BMT hypothesis gets better data to support it, at the time this paper was written, the conventional cladistics was actually following (unjustifiably) the hypothesis that was popular at the time – the verificationist approach was wrong, even if the results may happen to turn out right. Yet to the uninitiated like you and me, a cladogram in a Wikipedia article (or even, according to our authors, in the research literature) gives the impression of impressively supporting the hypothesis.
The fact that a better use of the methodology left the question far more open, even under the assumptions of evolutionary theory, should reinforce the thought with which we started: that cladistics can only give the best fit for any set of data one wishes to place within a nested hierarchy. Like my Russian dolls, it has no evidential value for the evolutionary significance of that nested hierarchy.
By way of rounding this off, I looked for more recent work that gives an idea of where the truth about bird phylogeny lies. As I have already mentioned, a new cladistic phylogeny seems still to put the origin of the birds in the theropoda, whilst making major changes to the postulated relationships of the theropods themselves. But I did find a 2015 paper that gives some evidence for some of the theropods being secondarily-flightless birds:
Was Caudipteryx derived from earth-bound theropod dinosaurs, which is the predominant view among palaeontologists, or was it secondarily flightless, with volant avians or theropods as ancestors (the neoflightless hypothesis), which is another popular, but minority view. The discovery here of an aerodynamic propatagium in several specimens provides new evidence that Caudipteryx (and hence oviraptorosaurs) represent secondarily derived flightless ground dwellers, whether of theropod or avian affinity, and that their presence and radiation during the Cretaceous may have been a factor in the apparent scarcity of many other large flightless birds during that period.
Caudipteryx
When I was doing my family history a couple a decade or so ago it was not uncommon to come across isolated pieces of contradictory evidence in parish records or a title deed suggesting that some individual was either the parent, or the child of some other individual – or maybe was actually two separate individuals. One chose one hypothesis over another on some fairly subjective criteria, not least because some person had to fill the gap in the genealogy. But in the nature of things one knew that, before the general resurrection of the dead enables the individuals concerned to set the record straight, the evidence will always be incomplete and disputible.
In the grand scheme of things one was also aware that it matters very little if two names on a list were identical in life, or alternatively entirely unrelated, even if one could prove it either way. When all one can do is provide a best fit for a conjectural family tree that is full of inexplicable anomalies anyway, nothing very much seems to hinge on it anyway.
“When all one can do is provide a best fit for a conjectural family tree that is full of inexplicable anomalies anyway, nothing very much seems to hinge on it anyway.”
Hey, are we back on Genesis genealogies again? Haha
Interesting post. Wish I knew more about cladistics, but my favorite part was the “ad hoc auxiliary hypothesis” anyway. Fun stuff!
Well Jay, cladistics is really just a way of doing classification, only unlike Linnaeus you don’t think in terms of genera, families etc, but the family tree (the clade) itself *is* the classification, as far as you wish to take it, or not. X nests within Y which nests within Z. You can sort out what to call it all later.
In theory you calculate it based on as many characters as possible – but of course when it comes to fossils, they’re mostly going to be skeletal traits, and in any case, like any classification system, you have to weight the traits in terms of what you deem their importance, especially when they start to generate anomalies.
I always think “Clade” is a good name for one of those gritty antiheroes, like “Klute”, with hi-tech gadgets like his Ad-Hoc Auxiliary Hypothesis.
“Hud” was my guy, another great name for an anti-hero. Paul Newman was reluctant to take the role, Hud was so incorrigible. I know too much about the movie because it was filmed 30 minutes from my hometown just after I was born. Not many movies filmed in the Texas Panhandle, unless they were documentaries about the Dust Bowl!
Yes, as far as weighing traits goes, I was just reading that the ultimate definition of “archaic” vs. “modern” H. sapiens might come down to our globular skull.
Looks like it’s time for the new, non-binary, incorrigible hard-person anti-hero, Florence. Bound to be a blockbuster in these confused times.
Yes, as far as weighing traits goes, I was just reading that the ultimate definition of “archaic” vs. “modern” H. sapiens might come down to our globular skull.
Just what I mean, Jay – whereas in reality it might have been more important in evolutionary terms that the archaic types used cold fusion rather than only fire, hadn’t hybridized with more primitive types etc.
More seriously, hybridization, like horizontal gene transfer, is one known mechanism that muddies the clear waters of clades. One can imagine a clade as a patrilinear genealogy: it’s suddenly less useful if one realises that people have mothers as well as fathers.
This yEC doesn’t agree with the classification we inherited from the middle ages.
I don’t agree there are such divisions in nature as dinosaurs.
So the stuff about birds being related to dinosaurs starts off with a flawed premise.
I don’t agree there are mammals or reptiles.
All they are ARE creatures, kinds, with some useful like traits from a creator.
they are not evidence of being a created division or of common descent.
Having mammary glands is just a good idea for totally unrelated kinds.
Therefore i suggest these bird like dinos, by way of looking at bones, really are just ground birds.
they are not dinos or reptiles at all.
Giant ground birds are a option for these “dinos’ whoo have very bird like bones.
A t rex didn’t roar but sang like a bird.
I speculate its a grand error.
Giant ground birds are a option for these “dinos’ whoo have very bird like bones.
Ah, but Robert, if you don’t believe in mammals or reptiles, why would you believe in birds?
The bible does have bird kinds and then the general bird type. Flying creatures very alike.
So ground birds is a accurate term. So my point is that since there were birds and birds with teeth then why not ground birds with teeth and a bit of muscle and other details.
So the idea of comparing dinos with birds in these cases is evidence the “dinos” were birds. Not birds are dinos but there are no dinos. Just kinds of creatures.
Everything in these matters is summing up traits and drawing conclusions about biological relationships.
Its not more sophisticated then that.
Mix and match. However there are presumptions here that I think are wrong and not proven.